Термин ретикулярная формация предложил в 1865 году немецкий ученый О. Дейтерс. Под этим термином Дейтерс понимал разбросанные в стволе головного мозга клетки, окруженные множеством волокон, идущих в различных направлениях. Именно сетевидное расположение волокон, связывающих между собой нервные клетки, послужило основой для предложенного названия.
В настоящее время морфологами и физиологами накоплен богатый материал о строении и функциях ретикулярной формации. Установлено, что структурные элементы ретикулярной формации локализуются в целом ряде мозговых образований, начиная с промежуточной зоны шейных сегментов спинного мозга (VII пластина), и заканчивая некоторыми структурами промежуточного мозга (интраламинарными ядрами, таламическим ретикулярным ядром). Ретикулярная формация состоит из значительного числа нервных клеток (в ней содержится почти 9/10 клеток всего ствола мозга). Общие черты строения ретикулярных структур - наличие особых ретикулярных нейронов и отличительный характер связей.
Рис. 1. Нейрон ретикулярной формации. Сагиттальный разрез ствола мозга крысенка.
На рисунке А представлен только один нейрон ретикулярной формации. Видно, что аксон разделяется на каудальный и ростральный сегменты, большой протяженности, со множеством коллатералей. Б. Коллатерали. Сагиттальный разрез нижней части ствола мозга крысенка, показывающий соединения коллатералей большого нисходящего пути (пирамидный путь) с ретикулярными нейронами. Коллатерали восходящих путей (сенсорные пути), отсутствующие на рисунке, соединяются с ретикулярными нейронами подобным же образом (по Шейбэлу М. Э. и Шейбэлу А. Б.)
Наряду с многочисленными отдельно лежащими нейронами, различными но форме и величине, в ретикулярной формации головного мозга имеются ядра. Рассеянные нейроны ретикулярной формации прежде всего играют важную роль в обеспечении сегментарных рефлексов, замыкающихся на уровне ствола головного мозга. Они выступают в качестве вставочных нейронов при осуществлении таких рефлекторных актов, как моргание, роговичный рефлекс и т. д.
Выяснено значение многих ядер ретикулярной формации. Так, ядра, расположенные в продолговатом мозге, имеют связи с вегетативными ядрами блуждающего и языкоглоточного нервов, симпатическими ядрами спинного мозга, они участвуют в регуляции сердечной деятельности, дыхания, тонуса сосудов, секреции желез и т. д.
Установлена роль голубого пятна и ядер шва в регуляции сна и бодрствования. Голубое пятно , находится в верхнелатеральной части ромбовидной ямки. Нейроны этого ядра продуцируют биологически активное вещество - норадреналин , который оказывает активирующее воздействие на нейроны вышележащих отделов мозга. Особенно высока активность нейронов голубого пятна во время бодрствования, во время глубокого сна она угасает почти полностью. Ядра шва располагаются по срединной линии продолговатого мозга. Нейроны этих ядер вырабатывают серотонин , который вызывает процессы разлитого торможения и состояние сна.
Ядра Кахаля и Даркшевича , относящиеся к ретикулярной формации среднего мозга, имеют связи, с ядрами III, IV, VI, VIII и XI пар черепных нервов. Они координируют работу этих нервных центров, что очень важно для обеспечения сочетанного поворота головы и глаз. Ретикулярная формация ствола головного мозга имеет важное значение в поддержании тонуса скелетной мускулатуры, посылая тонические импульсы на мотонейроны двигательных ядер черепных нервов и двигательных ядер передних рогов спинного мозга. В процессе эволюции из ретикулярной формации выделились такие самостоятельные образования, как красное ядро, черное вещество.
По структурно-функциональным критериям ретикулярная формация делится на 3 зоны:
1. Медианную, расположенную по средней линии;
2. Медиальную, занимающую медиальные отделы ствола;
3. Латеральную, нейроны которой лежат вблизи сенсорных образований.
Медианная зона представлена элементами шва, состоящие из ядер, нейроны которых синтезируют медиатор – серотонин. Система ядер шва принимает участие в организации агрессивного и полового поведения, в регуляции сна.
Медиальная (осевая) зона состоит из мелких нейронов, которые не ветвятся. В зоне располагается большое количество ядер. Встречаются также крупные мультиполярные нейроны с большим числом густо ветвящихся дендритов. Они образуют восходящие нервные волокна в кору больших полушарий и нисходящие нервные волокна в спинной мозг. Восходящие пути связи медиальной зоны оказывают активирующее влияние (прямо или опосредованно через таламус) на новую кору. Нисходящие пути оказывают тормозящее влияние.
Латеральная зона – к ней относятся ретикулярные образования, расположенные в стволе мозга вблизи сенсорных систем, а также ретикулярные нейроны, лежащие внутри сенсорных образований. Основным компонентом этой зоны является группы ядер, которые примыкают к ядру тройничного нерва. Все ядра латеральной зоны (за исключением ретикулярного латерального ядра продолговатого мозга) состоят из нейронов малой и средней величины и лишены крупных элементов. В этой зоне располагаются восходящие и нисходящие пути, обеспечивающие связь сенсорных образований с медиальной зоной ретикулярной формации и моторными ядрами ствола. Эта часть ретикулярной формации является более молодым и возможно прогрессивнее, с ее развитием связан факт уменьшения объема осевой ретикулярной формации в ходе эволюционного развития. Таким образом, латеральная зона – это совокупность элементарных интегративных единиц, сформированных вблизи и внутри специфических сенсорных систем.
Рис. 2. Ядра ретикулярной формации (РФ) (по: Niuwenhuys еt. аl, 1978).
 |
1-6 - медианная зона РФ: 1-4- ядра шва (1 - бледное, 2 - темное, 3 - большое, 4- мостовое), 5 - верхнее центральное, 6 - дорсальное ядро шва, 7-13 - медиальная зона РФ: 7 - ретикулярное парамедианное, 8 - гигантоклеточное, 9 - ретикулярное ядро покрышки моста, 10, 11 - каудальное (10) и оральное (11) ядра моста, 12 - дорсальное покрышечное ядро (Гуддена), 13 - клиновидное ядро, 14- I5-латеральная зона РФ: 14 - центральное ретикулярное ядро продолговатого мозга, 15 - латеральное ретикулярное ядро, 16, 17 - медиальное (16) и латеральное (17) парабрахиальные ядра, 18, 19 - компактная (18) и рассеянная (19) части педункуло-понтийного ядра.
Благодаря нисходящим влияниям ретикулярная формация оказывает тоническое влияние и на мотонейроны спинного мозга, что в свою очередь повышает тонус скелетной мускулатуры, совершенствует систему обратной афферентной связи. В результате любой двигательный акт совершается значительно эффективнее, осуществляет более точный контроль за движением, но чрезмерное возбуждение клеток ретикулярной формации может привести к дрожанию мышц.
В ядрах ретикулярной формации находятся центры сна и бодрствования, и стимуляция тех или иных центров приводит или к наступлению сна, или к пробуждению. На этом основано применение снотворных. В ретикулярной формации расположены нейроны, реагирующие на болевые раздражения, идущие от мышц или внутренних органов. В ней также расположены специальные нейроны, которые обеспечивают быструю реакцию на внезапные, неопределенные сигналы.
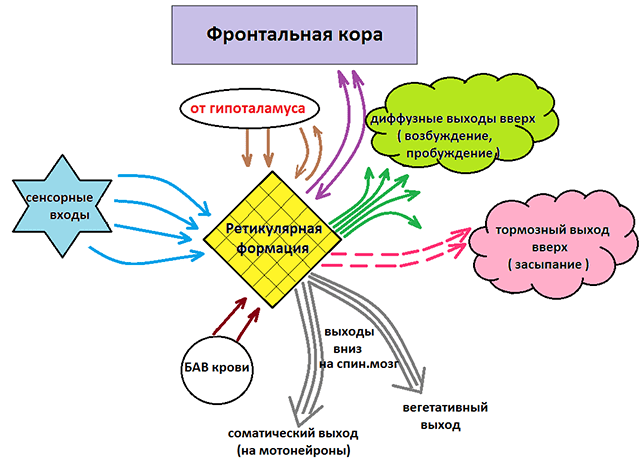
Ретикулярная формация тесно связана с корой больших полушарий, благодаря этому формируется функциональная связь между внешними отделами ЦНС и стволом головного мозга. Ретикулярная формация играет важную роль как в интеграции сенсорной информации, так и в контроле над деятельностью всех эффекторных нейронов (моторных и вегетативных). Она имеет также первостепенное значение для активации коры больших полушарий, для поддержания сознания.
Необходимо отметить, что кора полушарий большого мозга, и в свою очередь, посылает по корково-ретикулярным путям импульсы в ретикулярную формацию. Эти импульсы возникают в основном в коре лобной доли и проходят в составе пирамидных путей. Корково-ретикулярные связи оказывают либо тормозное, либо возбуждающее действие на ретикулярную формацию ствола головного мозга, они осуществляют корректировку прохождения импульсов по эфферентным путям (отбор эфферентной информации).
Таким образом, между ретикулярной формацией и корой полушарий большого мозга имеется двусторонняя связь, которая обеспечивает саморегуляцию в деятельности нервной системы. От функционального состояния ретикулярной формации зависит тонус мускулатуры, работа внутренних органов, настроение, концентрация внимания, память и т. д. В целом ретикулярная формация создает и поддерживает условия для осуществления сложной рефлекторной деятельности с участием коры полушарий большого мозга.
Сложное строение человеческого мозга открывает тайны нашего поведения, объясняет законы мыслительной деятельности, протекание эмоций и чувств. Каждое полушарие головного мозга отвечает за свои специфические функции и задачи (например, известно, что правое отвечает за логику, а левое - за воображение и фантазию), но имеются также структуры, которые обеспечивают единую и согласованную работу всей центральной нервной системы. Одной из таких структур является ретикулярная формация.
Общие сведения
Ретикулярная формация - это участок стволового отдела мозга, представленный разветвленной сетью нервных клеток и ядер, соединяющих между собой различные отделы головного мозга. В отличие от других структур, - например, таламуса, гипоталамуса, мозжечка, - которые имеют некую цельную форму (ядра, железы), ретикулярная формация не представлена единым морфологическим образованием, а является «сетью» (от латинского reticulum — сеть) дендритов и аксонов, которые с разной степенью плотности проникают между отделами и структурами головного мозга, объединяя их между собой и обеспечивая их совместную деятельность.
Метафорически выражаясь: если наш мозг представить в виде некоего изделия, скажем, рубашки, то ретикулярная формация - это нити, которыми рубашка сшита. Ретикулярная формация пронизывает структуры продолговатого, среднего мозга и моста, имеет непосредственные связи с мозжечком, спинным мозгом, таламусом и опосредованные - с вышележащими отделами: гипоталамусом, зрительными ядрами и корой.
Как устроена
В состав ретикулярной формации входит огромное количество нейронов с разветвленными дендритами и длинными аксонами, за счет чего становится возможным передавать нервные импульсы в различные отделы головного и спинного мозга. При этом можно выделить две наиболее крупные группы нейронных скоплений:
- Ретикулотегментальное ядро, нейроны которого получают сигналы от вышележащих отделов ГМ (четверохолмие, таламус) и передают их далее в структуры мозжечка, регулируя тем самым некоторые жизненно важные двигательные функции: координация взгляда, движения глаз.
- Латеральное ядро, нейроны которого восходят от структур спинного мозга и вестибулярных ядер и обеспечивают информирование коры ГМ о положении тела в пространстве, участвуют в регуляции дыхания и сосудистой иннервации.
- Кроме того, в состав ретикулярной формации входят нейроны, которые принимают важное участие в работе центров терморегуляции, насыщения и голода.

Основные функции
Основное предназначение ретикулярной формации состоит в сенсорном анализе многочисленных сигналов, поступающих от различных отделов ГМ.
За счет тесных связей со спинным мозгом она также принимает самое активное участие в моторной регуляции, начиная от глотательного рефлекса и заканчивая сложными двигательными операциями. Кроме того, ретикулярная формация осуществляет активирующее воздействие на весь ГМ в целом, участвуя в регуляции циклов сна и бодрствования.
В общем виде функции ретикулярной формации сводятся к следующим:
- Регуляция скелетной мускулатуры (участвует в управлении движениями тела) и вегетативных функций (дыхание, чихание, кровообращение и т.п.).
- Контроль процессов сна и бодрствования (за счет оказания активирующего и тормозящего воздействия на кору больших полушарий).
- Активизирующая функция (проявляется в том, что ретикулярная формация обеспечивает постоянное тоническое возбуждение коры ГМ, за счет чего становится возможным поддержание внимания, сознания и протекание мыслительных процессов.)
- Обработка сигналов, поступающих из внешней и внутренней среды.
Отличительные особенности работы ретикулярной формации связаны, прежде всего, с определенными свойствами ее :
Нервные клетки, входящие в состав ретикулярной формации, обладают повышенной способностью к тоническому возбуждению. Это означает, что бо́льшая часть нейронов находятся в постоянном возбуждении и генерирует нервные импульсы, которые передаются в вышележащие отделы ГМ. Такая тоническая активность обусловлена рядом факторов:
- Проходимость большого количества сигналов через структуры ретикулярной формации. Приведем простую аналогию: представьте себе рояль или какой-либо другой струнный инструмент. Понятно, что когда мы напрямую касаемся струн, они начинают колебаться и издавать звук. То же самое происходит и с нервными клетками, когда к ним приходят сигналы от других нейронов.Однако представим далее, что мы не касаемся непосредственно струн инструмента, но, скажем, скачем рядом с ним, сильно стуча ногами о пол. Звука инструмента мы, может быть, и не услышим, но едва заметное колебание струн все же будет иметь место. То же самое происходит и с нейронами ретикулярной формации. Поскольку через нее постоянно проходят какие-то сигналы (как афферентные, так и эфферентные) от различных структур ЦНС, то это и создает постоянное тоническое возбуждение нейронов ретикулярной формации, за счет того, что она находится в эпицентре постоянного нервно-импульсного обмена.
- Повышенная чувствительность нейронов к химическим веществам (гормоны, медикаменты, психотропные вещества). Чашка кофе, выпитая утром, «включает» именно структуры ретикулярной формации и за счет длительного сохранения возбуждения в ее нейронах поддерживает нас в активном состоянии.
Нисходящее и восходящее влияние РФ
Как уже отмечалось, ретикулярная формация оказывает возбуждающее и тормозящее действие на различные отделы ГМ. При этом можно выделить два отдела, которые специализируются на передаче возбуждения в те или иные структуры мозга.
Нисходящий отдел: представлен вегетативными и двигательными центрами и оказывает нисходящее влияние на отделы спинного мозга. Соответствующие нейронные скопления регулируют деятельность дыхательного, сосудодвигательного, слюноотделительного центров, а также центров, отвечающих за построение простых и сложных двигательных реакций. Это указывает на решающую роль именно центральной нервной системы в регуляции даже элементарных безусловных рефлексов. Стимуляция нисходящего отдела приводит к затормаживанию спинальных центров и вызывает в естественной среде состояние глубоко сна (спать «без задних ног»). Тот же эффект можно вызвать и искусственным путем, например, вводя человека в состояние транса или наркоза.
Восходящий отдел: представлен нервными волокнами, соединяющими структуры ретикулярной формации с вышележащими отделами: таламусом, гипоталамусом, мозжечком и корой. Восходящее влияние оказывает стимулирующее действие на кортикальные структуры и обеспечивает активное состояние сознания. Восходящее влияние не прекращается, даже когда мы спим. Если бы наш мозг мог полностью «отключаться», то каждое пробуждение было бы сродни рождению: кто я? Где я? Как я сюда попал? Однако за счет работы ретикулярных структур у нас сохраняется возможность всегда возвращаться в то исходное состояние сознания, в котором мы находились до момента сна. Кроме того, и во время ночного отдыха у нас остается способность реагировать на некоторые жизненно важные раздражители, т.е. мы, как правило, не спим «мертвым сном» и можем проснуться, если рядом зашевелился и заплакал ребенок, что-то громко упало и т.п.
Проявление повреждения структур
Ретикулярная формация играет значимую роль в интегративной деятельности всего головного мозга. За счет выполнения функции ведущего проводника всех видов нервных импульсов во все отделы ЦНС, ретикулярная формация находится в постоянной работе. Чрезмерные умственные и эмоциональные перегрузки вредны для мозга в целом и для ретикулярной формации, в частности. К счастью, своевременный прием седативных препаратов может (за счет повышенной восприимчивости нейронов к химическому воздействию) быстро исправить ситуацию и нормализовать состояние.
Однако возможны и менее благоприятные исходы. Повреждение возможно в результате черепно-мозговых травм, онкологических заболеваний головного мозга, инфекционных поражений.
Основным проявлением неблагополучия является потеря сознания.
Нарушение восходящих связей обнаруживает себя в состоянии апатии, слабости, повышенной сонливости, двигательной растороможенности, нарушениях ночного сна. Нередки сопутствующие вегетативные расстройства.
Впервые ретикулярная формация была описана в 1865 году немецким ученым О.Дейтерсом, который предложил и этот термин. Данным термином обозначали и продолжают обозначать участки мозга, в которых располагаются клетки различных размеров и форм, окруженные множеством волокон, идущих во всех направлениях. В конце прошлого века В.М.Бехтерев выделил в ретикулярной формации отдельные ядра.
Ретикулярная формация располагается в спинном мозге между задним и боковым рогами, а в ромбовидном и среднем мозге локализуется в покрышке. В ретикулярной формации ствола головного мозга человека описывают 22 ядра, которые объединяют в латеральную, медиальную и среднюю группы. Из среднего мозга ретикулярная формация продолжается в промежуточный мозг, в котором она представлена внутрипластинчатыми и ретикулярными ядрами таламуса. У низших животных ретикулярная формация имеет еще более широкое распространение в мозговом стволе. В процессе эволюции из нее выделяются как самостоятельные образования ядра моста, красное ядро, черное вещество.
Нервные связи ретикулярной формации весьма обширны. В ее латеральной трети находятся воспринимающие поля, в которых оканчиваются волокна различных афферентных систем, проходящих в стволе головного мозга. К ретикулярной формации подходят коллатерали медиальной и латеральной петель, часть чувствительных волокон V, VIII, IX и X черепных нервов. Медиальные 2/3 ретикулярной формации составляют эффекторные поля, связанные с двигательными ядрами черепных нервов, мозжечком, промежуточным мозгом, ядрами передних рогов спинного мозга.
Ядра ретикулярной формации, расположенные в продолговатом мозге, имеют связи с вегетативными ядрами блуждающего и языкоглоточного нервов, симпатическими ядрами спинного мозга. Поэтому они участвуют в регуляции сердечной деятельности, дыхания, тонуса сосудов, секреции желез и т.д.
Ядра Кахаля и Даркшевича, относящиеся к ретикулярной формации среднего мозга, имеют связи с ядрами III, IV, VI, VIII и XI пар черепных нервов. Они координируют работу этих нервных центров, что очень важно для обеспечения сочетанного поворота головы и глаз.
В составе ретикулярной формации имеются восходящие и нисходящие пути. Восходящие пути передают раздражения из нижних этажей ретикулярной формации к ядрам таламуса. Различают спинно-ретикулярный путь, начинающийся в ретикулярной формации спинного мозга; ретикуло-таламический путь, начинающийся в ретикулярной формации продолговатого мозга и моста; покрышечно-таламический путь, начинающийся в ретикулярной формации среднего мозга. Основным нисходящим путем является ретикулярно-спинномозговой путь, который берет начало в мосте и продолговатом мозге и идет к нейронам передних рогов спинного мозга и промежуточной части серого вещества. Посредством этого пути ретикулярная формация может оказывать облегчающее или тормозящее влияние на двигательные нейроны спинного мозга.
Верхние отделы ретикулярной формации связаны с корой большого мозга. Из внутрипластинчатых и ретикулярных ядер таламуса идут волокна в различные области коры. Признается также наличие прямых восходящих волокон из ретикулярной формации среднего мозга в кору. Особенностью ретикуло-корковых проекций является их диффузный характер, захватывающий все части коры. Этим они принципиально отличаются от специфических афферентных проекций различных видов чувствительности, которые связаны с определенными корковыми полями. Ретикуло-корковые волокна оканчиваются во всех слоях коры, тогда как местом окончания специфических чувствительных путей является внутренняя зернистая пластинка (IV слой коры).
Специфические и диффузная проекционные системы представляют собой 2 параллельных пути афферентных сигналов, направляемых в кору большого мозга. Различные раздражения, воспринимаемые рецепторами, передаются по специфическим афферентным системам в соответствующие воспринимающие области коры (общую чувствительную, зрительную, слуховую зоны и т.д.). Но вместе с этим в кору поступают неспецифические афферентные импульсы из ретикулярной формации по ее диффузной проекционной системе. Неспецифические импульсы осуществляют активацию коры, необходимую для восприятия специфических раздражений. Следует подчеркнуть важную роль неспецифических афферентных ретикулярных волокон в отборе (дифференцированном проведении импульсов) информации, поступающей к коре полушарий большого мозга. Прерывание потока импульсов из ретикулярной формации приводит к снижению тонуса коры, в результате чего наступает сон. Когда же прохождение импульсов из ретикулярной формации в кору восстанавливается, происходит пробуждение.
Установлена важная роль в регуляции сна и бодрствования относящихся к ретикулярной формации голубого пятна и ядер шва. Голубое пятно находится в верхнелатеральной части ромбовидной ямки, его нейроны продуцируют норадреналин, который по аксонам поступает в вышележащие отделы мозга. Активность этих нейронов максимальна во время бодрствования, снижается на ранних стадиях сна и почти полностью угасает во время глубокого сна. Ядра шва располагаются по средней линии продолговатого мозга. Нейроциты этих ядер вырабатывают серотонин, который вызывает процессы разлитого торможения и состояние сна.
Действие ретикулярной формации на кору большого мозга не является односторонним. Кора, в свою очередь, посылает в ретикулярную формацию сигналы. Эти сигналы идут по корково-ретикулярным волокнам, которые начинаются, в основном, в лобных долях полушарий и проходят в составе пирамидных путей к ретикулярной формации моста и продолговатого мозга. Корково-ретикулярные связи оказывают либо тормозное, либо возбуждающее действие на ретикулярную формацию ствола головного мозга, осуществляют корректировку прохождения импульсов по эфферентным путям (отбор эфферентной информации). Благодаря наличию двусторонней, кольцевой связи между ретикулярной формацией и корой может осуществляться саморегуляция корковой деятельности. От функционального состояния ретикулярной формации зависит тонус мускулатуры, работа внутренних органов, настроение, концентрация внимания, память и т.д. В целом ретикулярная формация создает и поддерживает условия осуществления сложной рефлекторной деятельности с участием коры полушарий большого мозга.
Несостоятельными оказались попытки представить ретикулярную формацию как высшую интегративную систему мозга, с которой будто бы связано сознание человека. В действительности сознание и умственная деятельность являются результатом постоянного взаимодействия коры и подкорковых центров, среди которых важная роль принадлежит ретикулярной формации. Как указывал академик П.К.Анохин, кора большого мозга выполняет в высшей нервной деятельности архитектурную функцию, а подкорковые образования обеспечивают ее энергетическую сторону.
Ретикулярная формация (РФ, сетчатый образование) - сердцевина мозга, представляет собой комплекс структур мозга, имеет большую протяженность. Начинается от желеобразной субстанции спинного мозга и заканчивается неспецифическими ядрами таламуса. Срок предложено Дейтерса. Клетки РФ, различной формы и размеров, имеют большое количество отростков, которые переплетаются между собой и образуют большое количество контактов (аксон в течение 2 см образует до 27000 синапсов). Под микроскопом РФ напоминает сетку, и стало основой для названия (лат. Reticularis - сетчатый). Нейроны сгруппированы в 48 отдельных ядрах, а также расположенные в составе многих структур ствола и промежуточного мозга.
Непосредственной связи с афферентными системами РФ не имеет, но к ней поступает коллатеральными путями вся чувствительная информация, направляется к таламуса.
Характерной особенностью нейронов ретикулярной формации является их постоянная фоновая активность, развивается вследствие:
1 высокой чувствительности клеток РФ к действию присутствующих или занесенных гуморальных агентов, постоянно недеполяризующие мембраны нейронов, возбуждают их;
2 огромного количества конвергентных контактов клеток РФ с коллатералями чувствительных нервных волокон, идущих к таламуса и постоянно раздражают ретикулярные нейроны.
В результате этого клетки РФ находятся в состоянии постоянной активности, которая распространяется на выше и ниже размещены структуры мозга. Распространение информации по мозговых структурах вверх и вниз дало возможность функционального разделения РФ на восходящую и нисходящую ретикулярные системы (рис. 4.25).
Функции нисходящей ретикулярной системы
Нисходящие влияния РФ представлены системой эфферентных нейронов, аксоны которых заканчиваются синапсами на нейронах структур мозга, расположенных ниже. В ее составе есть ядра, обеспечивающие регуляцию движений и мышечного тонуса .
Впервые влияние РФ на осуществление спинальных рефлексов было доказано И. М. Сеченова (1863) в опыте на "таламического" лягушке - открытии центрального торможения.
В середине XX века исследованиями Г. Мегуна было установлено, что ретикулярные ядра продолговатого мозга (гигантоклеточные) тормозят спинальные двигательные центры. В частности, было определено подавление децеребрации ригидности при раздражении ретикулярных ядер ствола мозга. Таким образом, только через 100 лет был раскрыт механизм сеченивського торможения: возбуждение ретикулярной формации через вставные тормозные нейроны (клетки Реншоу) вызывает торможение альфа-мотонейронов спинного мозга, в результате чего развивается неспецифическое торможение сгибательных и разгибательных спинномозговых рефлексов. Природа торможения альфа-мотонейронов связана с гиперполяризацией их мембран и соответствующего снижения их возбуждения. Кроме торможения, ретикулярная формация также вызывает и полегшувальний влияние на спинной мозг, центр которого находится в варолиева моста и среднем мозге (см. Рис. 4.25).
Кроме регуляции движений, ретикулярная формация влияет на ряд других функций. Известно, что в составе бульбарного дыхательного и кардиоваскулярного центров, слюноотделительного ядер, ядер блуждающего нерва и других находятся нейроны ретикулярной формации, которые активно участвуют в осуществлении жизненно важных функций организма - дыхания, кровообращения, пищеварения, теплообмена, обмена веществ.
Функции восходящей ретикулярной системы
Восходящий влияние ретикулярной формации на функцию коры мозга было впервые установлено Г. Мегуна
РИС. 4.25.

РИС. 4.26. Активирующие влияния ретикулярной формации и гипоталамуса на кору головного мозга
и Д. Моруцци (1949). Электрическое раздражение ретикулярных ядер ствола мозга с помощью хронически вживленных в них электродов воспроизводило в спящей кошки генерализованные изменения электрической активности коры мозга, характерные для активации, связанной с бодрствованием. Медленные волны в составе электроэнцефалограммы (ЭЭГ) превращались в быстрые, асинхронные. Эти изменения электрической активности мозга совпадали с реакцией пробуждения животного, его активным поведением (рис. 4.26). Это стало поводом для того, чтобы назвать восходящий влияние ретикулярной формации " восходящей активирующей системой ". К активирующих систем относятся и ретикулярные ядра среднего мозга - голубое пятно, которая активно участвует в строении сложных поведенческих реакций, внимания, памяти, обучения, эмоционально-мотивационных реакций. Однако при раздражении других частиц ретикулярной формации (варолиевого моста) могут развиваться тормозные эффекты.
Высокая чувствительность структур ретикулярной формации к различным гуморальных факторов свидетельствует о ее высокой надежности в регуляторных процессах организма, ее адаптивных реакций, направленных на поддержание гомеостаза.
Ретикулярная формация - совокупность различных , расположенных на протяжении ствола мозга, оказывающих активирующее или тормозящее влияние на различные структуры центральной нервной системы, тем самым контролируя их рефлекторную деятельность.
Ретикулярная формация ствола мозга оказывает активирующее влияние на клетки и тормозное действие на мотонейроны спинного мозга. Посылая в спинной мозг к его двигательным нейронам тормозящие и возбуждающие импульсы ретикулярная формация участвует в регуляции тонуса скелетных мышц.
Ретикулярная формация поддерживает тонус вегетативных центров, интегрирует симпатические и парасимпатические влияния, передает модулирующее влияние от гипоталамуса и мозжечка к внутренним органам.
Функции ретикулярной формации
Соматодвигательный контроль (активация скелетной мускулатуры), может быть прямым через tr. reticulospinalis и непрямым через , оливы, бугорки четверохолмия, красное ядро, черное вещество, полосатое тело, ядра таламуса и даже соматомоторные зоны коры.
Соматочувствительный контроль , т.е. снижение уровней соматосенсорной информации — «медленная боль», модификация восприятия различных видов сенсорной чувствительности (слуха, зрения, вестибуляции, обоняния).
Висцеромоторный контроль состояния сердечно-сосудистой, дыхательной систем, активности гладкой мускулатуры различных внутренних органов.
Нейроэндокринная трансдукция через влияние на нейромедиаторы, центры гипоталамуса и далее гипофиз.
Биоритмы через связи с гипоталамусом и шишковидной железой.
Различные функциональные состояния организма (сон, пробуждение, состояние сознания, поведение) осуществляются посредством многочисленных связей ядер ретикулярной формации со всеми частями ЦНС.
Координация работы разных центров ствола мозга , обеспечивающих сложные висцеральные рефлекторные ответы (чихание, кашель, рвота, зевота, жевание, сосание, глотание и др.).
Строение ретикулярной формации
Ретикулярная формация образована совокупностью многочисленных нейронов , лежащих отдельно или сгруппированных в ядра (см. рис. 1 и 2). Ее структуры локализуются в центральных участках ствола, начиная с верхних сегментов шейного отдела спинного мозга до верхнего уровня ствола мозга, где они постепенно сливаются с ядерными группами . Ретикулярная формация занимает пространства между ядрами черепных нервов, другими ядрами и трактами, проходящими через ствол мозга.
Нейроны ретикулярной формации характеризуются большим разнообразием форм и размеров, но их общим признаком является то, что они образуют длинными дендритами и широко ветвящимися аксонами многочисленные синаптические контакты как между собой, так и с нейронами других ядер мозга. Эти ветвления формируют своеобразную сеть (ретикулум ), откуда произошло название — ретикулярная формация. У нейронов, формирующих ядра ретикулярной формации, имеются длинные аксоны , образующие проводящие пути к спинному мозгу, ядрам ствола мозга, и других областей головного мозга.

Рис. 1. Важнейшие структурные образования среднего мозга (поперечный срез)
К нейронам ретикулярной формации поступают многочисленные афферентные сигналы из различных структур ЦНС. Можно выделить несколько групп нейронов, к которым поступают эти сигналы. Это группа нейронов латерального ядра ретикулярной формации, расположенного в продолговатом мозге. Нейроны ядра получают афферентные сигналы от вставочных нейронов спинного мозга и входят в состав одного из непрямых спиномозжечковых путей. Кроме того, они получают сигналы от вестибулярных ядер и могут интегрировать информацию о состоянии активности вставочных нейронов, связанных с мотонейронами спинного мозга, и о положении тела и головы в пространстве.
Следующая группа — это нейроны ретикулотегментального ядра , расположенные на границе дорсального края моста. Они получают афферентные синаптические входы от нейронов претектальных ядер и верхних холмиков четверохолмия и посылают свои аксоны в структуры мозжечка, участвующие в контроле движений глаз.
Нейроны ретикулярной формации получают разнообразные сигналы через пути, связывающие их с корой головного мозга (кортикоретикулоспинальные пути), черной субстанцией, и .

Рис. 2. Расположение некоторых ядер в стволе мозга и гипоталамусе: 1 — паравентрикулярное; 2 — дорсомедиальное: 3 — преоптическое; 4 — супраоптическое; 5 — заднее
Кроме описанных афферентных путей в ретикулярную формацию поступают сигналы по аксонным коллатералям проводящих путей сенсорных систем. При этом на один и тот же могут конвергировать сигналы от разных рецепторов (тактильных, зрительных, слуховых, вестибулярных, болевых, температурных, проприорецепторов, рецепторов внутренних органов).
Из приведенного перечня основных афферентных связей ретикулярной формации с другими областями ЦНС видно, что состояние ее тонической нейронной активности определяется притоком практически всех типов сенсорных сигналов от чувствительных нейронов, а также сигналов от большинства структур ЦНС.
Классификация ретикулярной формации в зависимости от направлении волокон|
Отделы |
Характеристика |
|
Нисходящий отдел |
Вегетативные центры:
Двигательные центры:
|
|
Восходящий отдел |
Ретикулоталамические Ретикулогипоталамические Ретикуломозжечковые Ретикулокортикальные: активирующие; гипногенные |
Ядра ретикулярной формации и их функции
Долгое время считалось, что ретикулярная формация, строение которой характеризуется широкими межнейронными связями, интегрирует сигналы различной модальности, не выделяя при этом специфической информации. Однако становится все более очевидным, что ретикулярная формация является не только морфологически, но и функционально гетерогенной, хотя различия между функциями ее отдельных частей не столь очевидны, как это характерно для других областей мозга.
Действительно, многие нейронные группы ретикулярной формации формируют ее ядра (центры), выполняющие специфические функции. Это нейронные группы, формирующие сосудодвигательный центр продолговатого мозга (гигантоклеточное, парамедианное, латеральное, вентральное, каудальное ядра продолговатого мозга), дыхательный центр (гигантоклеточное, мелкоклеточное ядра продолговатого мозга, оральное и каудальное ядра моста), центры жевания и глотания (латеральное, парамедианное ядра продолговатого мозга), центры движений глаз (парамедианная часть моста, ростральная часть среднего мозга), центры регуляции тонуса мышц (ростральное ядро моста и каудальное — продолговатого мозга) и др.
Одной из важнейших неспецифических функций ретикулярной формации является регуляция общей нейронной активности коры и других структур ЦНС. В ретикулярной формации проводится оценка биологической значимости поступающих сенсорных сигналов, и в зависимости от результатов этой оценки она может активировать или тормозить через неспецифические или специфические нейронные группы таламуса нейронные процессы во всей коре головного мозга или в се отдельных зонах. Поэтому стволовая ретикулярная формация называется также активирующей системой ствола мозга. Благодаря этим свойствам ретикулярная формация может оказывать влияние на уровень общей активности коры, поддержание которой является важнейшим условием для сохранения сознания, состояния бодрствования, формирования направленности внимания.
Повышение активности ретикулярной формации (на общем высоком фоне) в отдельных сенсорных, ассоциативных областях коры обеспечивает возможность выделения и обработки специфической, наиболее важной в данный момент времени информации для организма и организации адекватных ответных поведенческих реакций. Обычно эти реакции, организуемые при участии ретикулярной формации ствола мозга, предваряются ориентационными движениями глаз, головы и тела в направлении источника сигнала, изменениями дыхания и кровообращения.
Активирующее влияние ретикулярной формации на кору и другие структуры ЦНС осуществляется по восходящим путям, идущим от гигантоклеточного, латерального и вентрального ретикулярных ядер продолговатого мозга, а также от ядер моста и среднего мозга. По этим путям потоки нервных импульсов проводятся к нейронам неспецифических ядер таламуса и после их обработки переключаются в таламических ядрах для последующей передачи к коре. Кроме того, от перечисленных ретикулярных ядер потоки сигналов проводятся к нейронам заднего гипоталамуса и базальных ганглиев.
Кроме регуляции нейронной активности высших отделов мозга ретикулярная формация может регулировать сенсорные функции. Это осуществляется путем влияния на проведение афферентных сигналов в нервные центры, на возбудимость нейронов нервных центров, а также на чувствительность рецепторов. Повышение активности ретикулярной формации сопровождается повышением активности нейронов симпатической нервной системы, иннервирующей органы чувств. В результате может повышаться острота зрения, слуха, тактильная чувствительность.
Наряду с восходящими активирующими и тормозящими влияниями на высшие отделы головного мозга ретикулярная формация принимает участие в регуляции движений , оказывая активирующие и тормозные воздействия на спинной мозг. На ее ядрах происходит переключение как восходящих путей, идущих от проприорецепторов и спинного мозга к головному мозгу, так и нисходящих двигательных путей от коры мозга, базальных ядер, мозжечка и красного ядра. Хотя восходящие нейронные пути, идущие из ретикулярной формации в таламус и кору, играют роль преимущественно в поддержании общего уровня активности коры больших полушарий мозга, но именно эта их функция важна для осуществления бодрствующей корой планирования, запуска, исполнения движений и контроля за их исполнением. Между восходящими и нисходящими через ретикулярную формацию путями имеется большое число коллатеральных связей, через которые они могут оказывать взаимное влияние. Существование такого тесного взаимодействия создает условия для взаимного воздействия области ретикулярной формации, влияющей через таламус на активность коры, планирующей и инициирующей движения, и области ретикулярной формации, влияющей на исполнительные нейронные механизмы спинного мозга. В ретикулярной формации имеются группы нейронов, которые посылают большинство аксонов в мозжечок, участвующий в регуляции и координации сложных движений.
По нисходящим ретикулоспинальным путям ретикулярная формация непосредственно воздействует на функции спинного мозга. Прямое влияние на его двигательные центры осуществляется по медиальному ретикулоспинальному тракту, идущему от ядер моста и активирующему преимущественно интер- и у-моторные нейроны разгибателей и тормозящему моторные нейроны мышц-сгибателей туловища и конечностей. По латеральному ретикулоспинальному тракту , начинающемуся от гигантоклеточного ядра продолговатого мозга, ретикулярная формация оказывает активирующее влияние на интер- и у-моторные нейроны мышц-сгибателей конечностей и тормозящее на нейроны мышц-разгибателей.
Из экспериментальных наблюдений на животных известно, что стимуляция более рострально расположенных нейронов ретикулярной формации на уровне продолговатого и среднего мозга, оказывает диффузное облегчающее влияние на спинальные рефлексы, а стимуляция нейронов каудальной части продолговатого мозга сопровождается торможением сиинальных рефлексов.
Активирующее и тормозное влияние ретикулярной формации на моторные центры спинного мозга может осуществляться через у-мотонейроны. При этом ретикулярные нейроны рострального участка ретикулярной формации активируют у-мотонейроны, которые своими аксонами иннервируют интрафузальные мышечные волокна, вызывают их сокращение, активируют рецепторы мышечных веретен. Поток сигналов от этих рецепторов активирует а-мотонейроны и вызывает сокращение соответствующей мышцы. Нейроны каудального участка ретикулярной формации тормозят активность у-мотонейронов спинного мозга и вызывают расслабление мышц. От баланса нейронной активности этих участков ретикулярной формации зависит распределение тонуса в больших мышечных группах. Поскольку этот баланс зависит от нисходящих влияний на ретикулярную формацию коры головного мозга, базальных ганглиев, гипоталамуса, мозжечка, то эти структуры мозга также могут через ретикулярную формацию и другие ядра ствола мозга воздействовать на распределение тонуса мышц и позу тела.
Широкое ветвление аксонов ретикулоспинальных путей в спинном мозге создает условия для влияния ретикулярной формации практически на все моторные нейроны и соответственно на состояние мышц различных частей тела. Такая особенность обеспечивает эффективное воздействие ретикулярной формации на рефлекторное распределение тонуса мышц, позу, ориентацию головы и тела в направлении действия внешних раздражителей и участие ретикулярной формации в осуществлении произвольных движений мышц проксимальных частей тела.
В центральной части ретикулярного гигантоклеточного ядра располагается участок, раздражение которого тормозит все двигательные рефлексы спинного мозга. Наличие такого торможения структур головного мозга на спинной мозг было открыто И.М. Сеченовым в опытах на лягушках. Суть опытов состояла в исследовании состояния рефлексов спинного мозга после перересечения ствола мозга на уровне промежуточного мозга и раздражении каудального участка разреза кристалликом поваренной соли. Оказалось, что двигательные спинальные рефлексы при раздражении не проявлялись или становились ослабленными и восстанавливались после устранения раздражения. Таким образом было впервые выявлено, что один нервный центр может тормозить активность другого. Это явление назвали центральным торможением.
Ретикулярная формация играет важную роль в регуляции не только соматических, но и вегетативных функций (ретикулярные ядра ствола мозга входят в структуру жизненно важных отделов дыхательного центра и центров регуляции кровообращения). Латеральная группа ретикулярных ядер моста и дорсолатеральное ядро покрышки формируют мочевыделительный центр моста. Аксоны нейронов ядер этого центра достигают преганглионарных нейронов крестцового отдела спинного мозга. Стимуляция нейронов этих ядер в мосту сопровождается сокращением мускулатуры стенки мочевого пузыря и мочевыделением.
В дорсолатеральном мосту расположено парабрахиальное ядро, на нейронах которого заканчиваются волокна сенсорных нейронов вкуса. Нейроны ядра, подобно нейронам голубоватого пятна и черной субстанции, содержат нейромеланин. Число таких нейронов в парабрахиальном ядре уменьшается при болезни Паркинсона. Нейроны парабрахиального ядра имеют связи с нейронами гипоталамуса, амигдалы, ядрами шва, одиночного тракта и другими ядрами ствола мозга. Предполагают, что парабрахиальные ядра имеют отношение к регуляции вегетативных функций и понижение их числа при паркинсонизме объясняет возникновение вегетативных нарушений при этом заболевании.
В опытах на животных было показано, что при раздражении некоторых локальных участков ретикулярных структур продолговатого мозга и моста можно вызвать торможение активности коры и сон. На ЭЭГ при этом возникают низкочастотные (1-4 Гц) волны. На основе описанных фактов считают, что важнейшими функциями восходящих влияний ретикулярной формации являются регуляция цикла сон — бодрствование и уровня сознания. Оказалось, что к формированию этих состояний имеют прямое отношение ряд ядер ретикулярной формации ствола мозга.
Так, с каждой стороны центрального шва моста располагаются парамедианные ретикулярные ядра, или ядра шва , содержащие серотонинергические нейроны. В каудальной части моста они включают нижнее центральное ядро, которое является продолжением ядра шва продолговатого мозга, а в ростральной части моста в состав ядер шва моста входит верхнее центральное ядро, называемое ядром Бехтерева, или срединным ядром шва.
В ростральной части моста на дорсальной стороне покрышки располагается группа ядер голубоватого пятна. В них имеется около 16 000-18 000 меланинсодержащих норадренергических нейронов, аксоны которых широко представлены в различных отделах ЦНС — гипоталамусе, гиппокампс, коре больших полушарий мозга, мозжечке и спинном мозге. Голубоватое пятно простирается в средний мозг, и его нейроны прослеживаются в ссром веществе околоводопроводного пространства. Число нейронов в ядрах голубоватого пятна уменьшается при паркинсонизме, болезни Альцгеймера и синдроме Дауна.
Как серотонинергические, так и норадреналинергические нейроны ретикулярной формации играют роль в контроле цикла сон — бодрствование. Подавление синтеза серотонина в ядрах шва ведет к развитию бессонницы. Предполагают, что серотонинергические нейроны являются частью нервной сети регуляции медленноволнового сна. При действии серотонина на нейроны голубоватого пятна возникает парадоксальный сон. Разрушение ядер голубоватого пятна у экспериментальных животных не ведет к развитию бессонницы, но вызывает на несколько недель исчезновение фазы парадоксального сна.







